
I’ve been doing some research on the origin of body plans. Mainly to address those chapters in Darwin’s Doubt that deal with them. Nick Matzke put me on the trail of some recent work in the area. While reading the article (published in 2009), I saw some compelling evidence that the body plans of the various phyla developed prior to the Cambrian explosion. If this were true, then the entire point of Meyer’s book is refuted.
If animals had the genes for the body plans and only something like the regulation of those genes changed, then Meyer’s various claims about body plans suddenly appearing in the Cambrian and no evidence for prior forms are destroyed. I’ll get to that evidence in a minute.
As I was reading this article, I wondered if Meyer had read it. Turns out, he does actually quote from the article, but it is only included in a footnote, not in the references. This omission is interesting when we see just what Meyer says… and doesn’t say about the article.
The article in question is by Charles Marshall and James Valentine, both victims of massive quotemining by Meyer in this book. The article is titled “The Importance of Preadapted Genomes in the Origin of the Animal Bodyplans and the Cambrian Explosion“. Wow, not only is it a mouthful, but something like this should be a primary work cited in a book that concerns itself with the origin of body plans, the genetics of them, and the Cambrian Explosion. But it’s just a little, unimportant footnote. I wonder why Meyer is hiding this… let’s find out.
When trying to unravel the origins of the animal phyla… the hardest to examine is the phase between the actual cladogenic origin of a phylum and the time that it acquired its first phylum-specific characteristic(s). Even if we have fossils from this phase in a phylum’s history, we will not be able to prove their kinships at the level of phyla.54 [ed note: Darwin’s Doubt pg.95]
This is what Meyer wrote. The “54” is the footnote. And there, are those damn ellipses again. Let’s see what’s missing, by looking at what the actual article says.
When trying to unravel the origins of the animal phyla, three distinct phases in their evolution must be considered (Fig. 1). First, and the hardest to examine, is the phase between the actual cladogenic origin of a phylum, and the time that it acquired its first phylum-specific characteristic(s). Even if we have fossils from this phase in a phylum’s history, we will not be able to prove their kinships at the level of phyla, although they may be referable to a more inclusive clade within which the phylum has branched. [ed note: I bolded the differences.]
Wow, that’s a pretty blatant quotemine there. Notice how Meyer (through the use of those magical ellipses) removed the entire idea that there are three distinct phases AND the idea that phylums can be associated with a clade at all.
In other words, Meyer took one part of the ideas presented by Marshall and Valentine and created from it the idea of an insurmountable problem, when in reality it’s not insurmountable and it’s not really a major issue at all.
What do the authors say about the other two pieces that Meyer ignored?
In contrast, phase 2 consists of the stem lineages that descended from the first species that can be diagnosed morphologically as belonging to the phylum. However, some of the morphological features that define a phylum may have low preservation potentials, so that even if we have early fossils from this phase of a phylum’s evolution, we may not see the key features that would permit accurate assignment. Thus, as a practical paleontological matter, even if we are lucky enough to find early fossils that just postdate the origin of the first morphologically diagnostic feature of the phylum, they may not be identified as such.
This is really the problem of fossilization that I’ve talked about earlier. We’re talking about fossils that have survived over half a billion (with a “B”) years of Earth’s changes. The changes are major changes to entire continents and oceans.
When we find the fossils and when they have parts that fossilize well (chitin, for example), then we can (usually) put the organism into a phylum. But if we don’t have hard parts (things like the cell structure, or DNA, or muscles), then we may not even realize that an organism really should belong into a particular phylum. For example, it wasn’t until recently that significant evidence turned up that birds really did come from dinosaurs. And that was only about 100 millions years ago, not 500 million years ago.
Phase 3 begins with the appearance of the last common ancestor of the living representatives of the phylum, that is, of the crown group of the phylum. Here the fossil record, as well as comparative analysis of the living taxa, informs our understanding of the morphology of this LCA of the crown group.
Here, we get the recognizable phyla and can easily determine the existence of the characters that define the phyla.
The authors state that the purpose of the article is to deal with phase 1. Meyer ignores the remainder of the 9 pages of the article, which contains ideas and evidence about where the body plans of the phyla came from. Let me just post some quotes from the article. Obviously, if Meyer considers the work good enough to put in his book, then he agrees that Marshall and Valentine are experts right… and I think it’s important to let them have their say.
To examine this part of the animal tree, the fossil record is of little use (but see below). Instead we must rely heavily on both the morphology and on molecular characters from living taxa to determine the topology of the tree, and then, having secured that, turn to comparative genomic data to make inferences about the developmental and morphological and ecological capacities of those now extinct basal taxa (Table 2).
Inferences are good enough to the notions of Intelligent Design right? But these inferences will have statistical support.
Here we are interested in the relations between those deeper-branching phyla for which whole genomes or significant amounts of data on developmental regulatory genes are available. Of all the topologies that have been published, we feel that the topology shown in Figure 2 is the most likely, and each node finds support from both morphological and molecular data. This topology implies an increase in the size of the regulatory gene complement from branch to branch, indicated by the increasing numbers of regulatory gene families found, as reviewed below.
I have added figure 2 below.
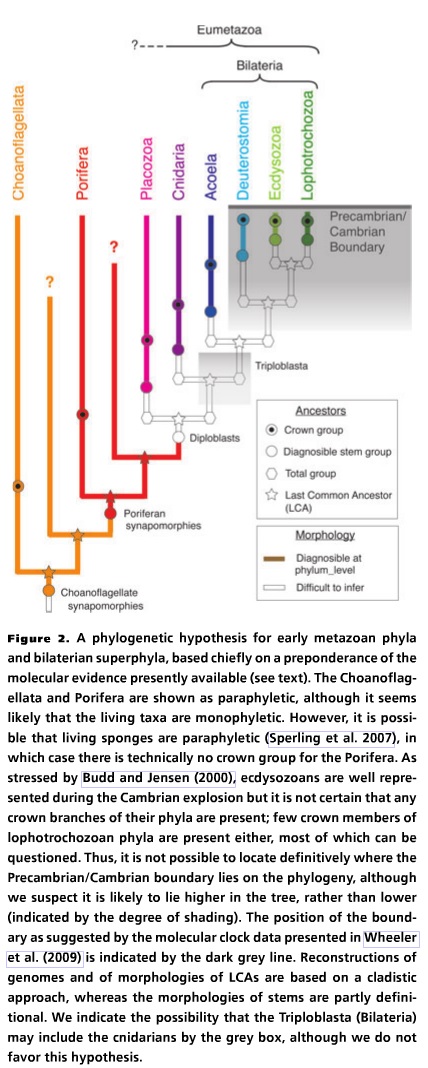
Just so you can see what their reconstruction looks like. The important bit is the statement about how the number of regulatory genes increase from branch to branch. What the authors are saying is that, unlike the high school level biology that most creationists (and non-specialists) have where DNA makes proteins and that’s it. There are lots and lots of genes that don’t create proteins. These DNA sequences regulate other DNA sequences and some of those DNA sequences produce proteins. I wrote about this earlier.
Clearly, the pre [Cambrian] explosion fauna had to have been phylogenetically diverse, as it included the ancestors of the explosion phyla. Furthermore, the position of explosion fossils on the phylogenetic tree implies that many phyla that do not appear during the explosion were nevertheless present at least as stems (phases 1 and 2). Features associated with larger bodyplans (and not required of tiny organisms) were presumably elaborated as the larger forms evolved, producing the beautiful stem forms of the explosion fauna.
What they are saying (with a page and half of data and references), is that the preCambrian animals were very diverse. The fossils from the Cambrian imply that the phyla that appeared after the Cambrian were present, just in a form we don’t realize belong to that phyla (phases 1 and 2 mentioned above).
Far from being a sudden appearance of all the phyla, they were either A) already present, we just can’t tell or B) the ancestors that became those phyla were already present. The data presented here (which I haven’t found and read all of yet, there are over two pages of references) suggests that Meyer is totally wrong. And his failure to at least include this work (and those referenced by this work) in his book is a scholarly failure of the highest order.
When taken at face value, the message from all these data is that many of the genes that regulate eumetazoan and bilaterian development originated in relatively simple animals before and during the origin of eumetazoans, before they acquired their well known functions in bilaterians, functions commonly conserved across many phyla. Certainly the accumulation in early metazoan genomes of what are today important regulatory genes does not seem to correlate with morphological complexity; that is, the early metazoans are much simpler than one might suspect give those gene functions in more derived metazoans. But perhaps any such expectation is simply over-interpreting early functions of those genes; it seems likely that although they serve in the development
of morphologically complex organisms now, their presence does not mean that the organisms that possessed them were necessarily complex.
This statement follows another page or so of data and references. If you can read through all the odd names, this breaks down to the genes that are important bilateralians (animals with a left and a right side, like humans) are present well before the first bilateralian existed. Metaazoans are animals. The early metazoans were simple organisms, but they already had the genes that were needed to make more complex organisms.
It seems reasonable to suggest that the Cambrian explosion in bilaterian body plans was underlain chiefly by selection for novel patterns of gene expression, including regulatory RNAs as well (see below), mediating the morphological radiation as the bilaterian lineages responded to new adaptive opportunities.
I am reminded of Ghostbusters here. I can see this little organism saying “We have the tools, and we have the talent.” We need to get away from (again) this high school biology idea of an organism (or population) changing in response to a stimulus. Evolution isn’t “It’s getting hotter, we have to adapt.” Evolution is, “this is what we have, what can we do with it”. Provided we anthropomorphicize evolution, which we shouldn’t do, because it’s wrong.
We see, through the data that Marshall and Valentine provide that the genes were already in place. Maybe they weren’t yet doing what they do in modern organisms, but they were there. How do we know that?
Some genes are present in every single animal on the planet. Genes like the Hox genes. It is much more likely that these genes evolved once, in the last common ancestor of all animals, than evolved almost exactly the same, many times, in many different phyla. Now, a design advocate would say, “It’ not common ancestry, but common design.”
That’s fine as far as it goes, but all that claim is doing is pawning off the issue until the designer is found. Until ID advocates find the designer, then that argument is essentially useless.
Given that the genomes of the many invertebrate lineages that were ancestral to the disparate stem and crown groups of the explosion fauna harbored rich developmental genomes (as is certain regardless of whether they were reduced or cryptic), it is easier to understand the relatively abrupt evolution of the bodyplans of larger, more complex explosion fossils. The requirements for increased developmental complexity and refinement needed to generate those morphological novelties did not have to be met from scratch; by and large the required pool of regulatory genes was already present in the more basal bilaterian lineages and the mechanisms of signaling and transcription pattern evolution were already well established, even if they were later elaborated through gene duplication in some lineages.
Again, the authors are just stating what we’ve already talked about. The genes for complexity were already present. Unlike what Meyer claims, the genes needed to create these body plans didn’t have to ‘suddenly appear’, but just minor regulatory changes and you get massive changes in the appearance of the organisms. Just as humans have individuals ranging from 3 foot tall to almost 9 feet tall… and there weren’t massive changes to the entire genetic code to generate those differences.
This paper is a really amazing piece of work and I’m still digesting it. I’ll probably do more writing on it later. If I haven’t been clear, please let me know, this was kind of a rush writing job (and I detest editing myself… as you can probably tell).
In conclusion, Meyer, once again, takes a quote out of context and mangles it in the hopes that no one will call him on it. Sorry buddy, but I’m calling you on it. There are two pages of references that support the work in this article… when you have refuted the majority of them with data… well, even then your quotemine isn’t justified.
____________________________
Marshall, Charles , and James Valentine. “The importance of preadapted genomes in the origin of the animal bodyplans and the Cambrian explosion.”Evolution; international journal of organic evolution, 30 Apr. 2010. Available: http://www.ncbi.nlm.nih.gov/pubmed/19930449.